
Prof. dr hab. Stanisław Wężyk
Dr inż. Ryszard Gilewski
W ostatnich dziesięcioleciach w produkcji drobiarskiej nastąpił ogromny postęp genetyczny, kosztem często dużych nakładów związanych np. z genotypowaniem ptaków tak by nawet niewielki zysk genetyczny na poziomie czystych linii, był zwielokrotniony w produkcji towarowej.

Genotypowanie jest określaniem lub sprawdzaniem molekularnymi technikami np. odczytywaniem sekwencji, czyli kolejności par nukleotydowych w cząsteczce DNA (genów lub ich fragmentów), lub zmian w tych rejonach (mutacji, polimorfizmów) celem określania poszczególnych genotypów. W tym kontekście niezwykle ważną jest precyzja oceny potencjału genetycznego poszczególnych ptaków, czyli ich genomu.
Genom, która to nazwa pochodzi z połączenia słów gen i chromosom, stanowi kompletną, generyczną informację o żywym organizmie lub wirusie. W przypadku organizmów eukariotycznych (jądrowych) termin ten odnosi się zwykle do materiału genetycznego zawartego w podstawowym, pojedynczym (haploidalnym) zespole chromosomów.
Organizm roślinny i zwierzęcy zbudowany jest z komórek z jądrem komórkowym zawierającym chromosomy otoczone błoną jądrową, w których DNA powiązany jest z białkami, tworząc chromatynę. Komórki eukariotyczne wyposażone są też w odrębne organelle komórkowe, takie jak mitochondria czy plastydy, które zawierają materiał genetyczny, kierując procesami metabolicznymi/życiowymi.
Ocena użytkowości obejmuje coraz liczebniejsze grupy cech i lawinowo rosnącą ilość informacji molekularnych dotyczącą SNP-ów, ekspresję genów i ich modyfikacji, oraz coraz większe wymagania stawiane badaniom łączących metody hodowli drobiu, genetykę, weterynarię, statystykę i bioinformatykę (Szwaczkowski, 2015). Polimorfizm pojedynczego nukleotydu (ang. Single Nucleotide Polymorphism w skrócie SNP) odzwierciedla zmienność sekwencji DNA, polegającej na wymianie pojedynczego nukleotydu (A, T, C lub G) pomiędzy osobnikami lub chromosomami danego osobnika. W programach hodowli drobiu, schematy w kształcie piramidy są zwykle stosowane w produkcji mieszańców pochodzących z kilku linii czystych, z których każda jest selekcjonowana niezależnie od pozostałych, a jej genetyczną ocenę przeprowadza się na podstawie wyników produkcyjnych ptaków czystorasowych, które nie są doskonałymi, przewidywalnymi wskaźnikami wyników produkcyjnych towarowych mieszańców. Nieaddytywne efekty genetyczne w postaci dominacji i epistazy lub genotypu warunkującego dla danej cechy interakcje środowiskowe i korelacje genetyczne między ptakami czystorasowymi a mieszańcami – mogą znacznie się różnić. By ominąć te ograniczenia, Wei (1992) zaproponował połączenie informacji uzyskanych od ptaków czystorasowych oraz mieszańców i wykorzystać je w metodach produkcji mieszańców towarowych. W praktyce metoda ta ogranicza się tylko do genotypowania ptaków czystorasowych (Picard Druet in., 2019). Z kolei Christensen i in. (2014), rozszerzyli model Wei i van der Werf’a (1994) stosując jednoetapową metodę, pozwalającą na wspólną ocenę ptaków genotypowanych lub nie genotypowanych. Powszechnie stosowana przez firmy hodowlane kur nieśnych strategia polega na sprawdzaniu ogólnej zdolności wykorzystania czystorasowych kogutów, na podstawie wyników produkcyjnych kur z nimi w różnym stopniu spokrewnionych. Druet i in., (2019), ocenili metodą BLUP Najlepsza Liniowa Nieobciążona Predykcja w czystorasowej linii kur genom cech jakości jaj, wykorzystując wyniki kur-mieszańców pochodzących z tej samej linii. Predykcja, czyli prognozowanie określenie stosowane we współczesnej genomice, jest racjonalnym, naukowym przewidywaniem przyszłych zdarzeń. Dla nadchodzącego okresu jest wyborem najbardziej prawdopodobnego rozwoju zjawiska, którego celem jest zmniejszenie ryzyka w podejmowaniu decyzji.
Materiał i metoda oceny genomu kur
Ptaki i genotypy
Druet i in. (2019), objęli badaniami 7982 czystorasowe kury pochodzące po 514 kogutach-ojcach i 1759 kurach-matkach rasy Rhode Island hodowli firmy Novogen, wylęgnięte w latach 2008 – 2015. Cechy jakości jaj mierzono raz w tygodniu, od 60 do 80 tygodnia życia kur, które oceniano, uwzględniając w ocenie 580.961 markerów SNP. Genotypowaniem SNP objęto zmiany genetycznych polimorfizmów pojedynczych nukleotydów (SNP) między osobnikami tego samego gatunku będącymi najczęstszymi rodzajami zmian
genetycznych.
Po określeniu 302102 genotypów SNP, 1214 kogutów i 1148 kur poddano dalszymi badaniami. Oceniono także produkcyjność mieszańców, których ojcami były czystorasowe koguty wylęgnięte w trzech pokoleniach.
Każdy czystorasowy kogut-ojciec miał 45 córek mieszańców, z nieznanymi kurami-matkami, o różnym genotypie. Kury mieszańce utrzymywano w klatkach po 12 przyrodnich sióstr pochodzących po tym samym ojcu. Jakościowe cechy jaj mierzono po osiągnięciu przez nioski 70 tygodni życia.
Cechy jakości jaj
Badano następujące cechy jakości jaj: masę jaja (MJ), kolor skorupy (KS) i jej wytrzymałość (WS). Oceniano masę poszczególnych jaj (w g), kolor ich skorupy chromometrem Minolta. Elastyczne odkształcenie i wytrzymałość skorupy jaja, mierzono określając jej wytrzymałość, poprzez ściskanie jaja między dwoma płytkami. Wielkość WS odpowiadała maksymalnej sile zarejestrowanej tuż przed pęknięciem skorupy jaja.
Ocena genetyczna i genomowa
Przed przystąpieniem do oceny, wartości cech uśredniono i określając ich standard. Każdą cechę najpierw oceniono na podstawie wydajności kur czysto rasowych, stosując jedno-cechowy model zwierzęcia, a potem dwu-cechowe modele zwierzęcia.
Dla wszystkich cech model statystyczny uwzględniał wpływ środowiska, usytuowanie klatki w kurniku, czas oczekiwania pomiędzy pobraniem próbki a pomiarem cechy jaja (w dniach) uwzględniając wiek kury (w tygodniach).
W przypadku kur czystorasowych przeprowadzano kilka pomiarów, uwzględniając w modelu losowym, wspólny wpływ środowiska na kury.
Ocenę przeprowadzono, wykorzystując metodę BLUP (dla oszacowania wartości hodowlanej) oraz jednoetapową metodę GBLUP w celu określenia Genomowej Wartości Hodowlanej (GWH), wdrożonej w rodzinowych programach, w celu analizy dokładności genomowego najlepszego, liniowego, nieobciążonego prognozowania wartości hodowanej.
Przewidywanie niezawodności oceny
Aby ocenić przydatność ocen, oszacowane wartości genetyczne i genomowe porównano z ich rzeczywistymi genetycznymi wartościami, badając czystorasowe 93 koguty i kury-córki oraz córki mieszańce, wykorzystując informacje o fenotypach ocenianych świeżo wyklutych piskląt i gdy osiągnęły one wiek 80 tygodni.
W pierwszym przypadku fenotypowanie populacji było ograniczone do przodków ocenianych piskląt, a w drugim – fenotypowana populacja obejmowała zarówno przodków, jak i współczesnych krewnych ptaków objętych programem hodowlanym. Przykład ten jest klasycznym schematem doboru kur nieśnych. Dostępne fenotypy stanowiły kompletny zbiór danych, który zawierał wyniki kur – córek czystorasowych i mieszańców.
Omówienie wyników
Parametry genetyczne
Wartość genetyczną określaną metodą BLUP, gdzie model zwierzęcia wyrażony jest liczbowo wartością odchylenia standardowego od średniej wartości cech wszystkich osobników, uwzględnionych w obliczeniach. Końcowym wynikiem jest ranking ptaków na podstawie liczbowych wartości. Metoda ta pozwala oszacować faktyczną wartość genetyczną ptaków oraz podjąć decyzje hodowlane, nim zostanie w pełni poznana ich wartość użytkowa.
Genomowe najlepsze liniowe, nieobciążone prognozowanie (GBLUP) jest metodą, która wykorzystuje zależności genomowe do oszacowania wartości genetycznej osobnika.
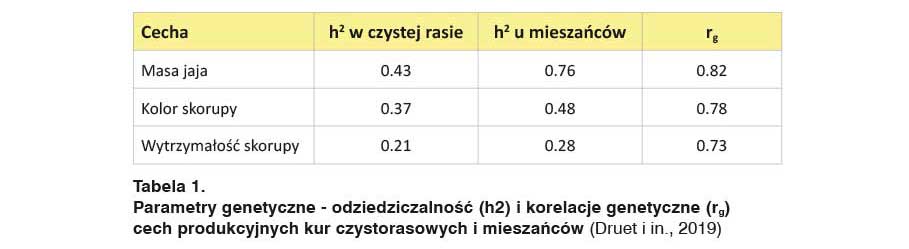
Oszacowane współczynniki odziedziczalności (h2) w populacji czystorasowej były zgodne z przytaczanymi w literaturze. Były one wyższe niż w populacji mieszańców, prawdopodobnie na skutek obecności nieaddytywnych efektów genetycznych – szczególnie w przypadku masy jaja.
W przypadku wszystkich cech zaobserwowano silny związek między cechami ptaków czystorasowych i mieszańców, z korelacjami genetycznymi o wartościach rg=0,73 – 0,82 (Tab. 1).
Wartości te znacznie różniły się od „1”, co wskazuje na uzasadnione zainteresowanie dwuzmiennymi modelami, wykorzystywanymi do dokładnego szacowania genomowej wartości hodowlanej, zarówno dla cech w populacjach czystorasowych, jak i u mieszańców towarowych drobiu.
Względna dokładność oceny wartości hodowlanej i genomowej drobiu
W przypadku wszystkich cech i ocen, względna dokładność oceny genomowej znacząco wzrosła między oceną przeprowadzoną przy wykluciu, a oceną jaj przeprowadzoną, gdy kury miały 80 tygodni życia (Tabela 2).
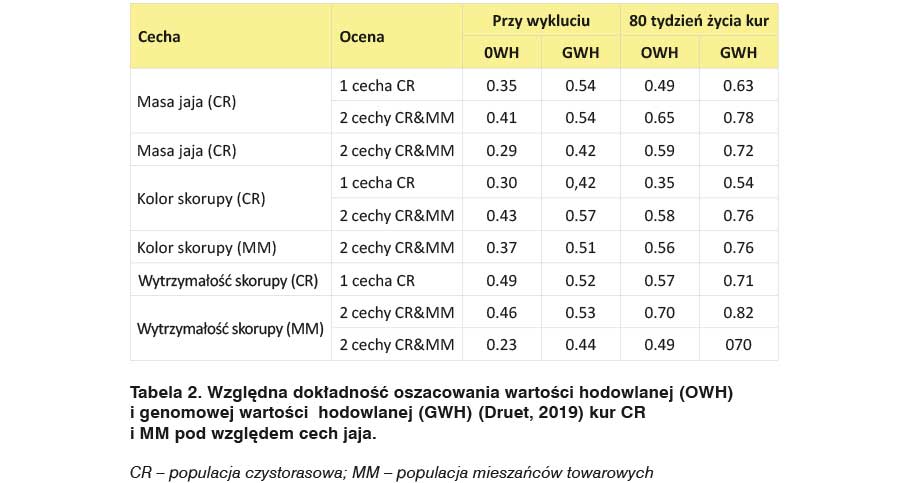
W przypadku modeli dwucechowych przyrost dokładności między oceną hodowlaną, a genomową był zawsze znaczący, chociaż różny w zależności od cechy i okresu prowadzonej oceny.
Odchylenie między cechami w populacji czystorasowej i mieszańców było umiarkowane w odniesieniu do masy jaja i koloru skorupy. Było większe w przypadku wytrzymałości skorupy i wahało się od 13% w czystej rasie do 48% przy kluciu mieszańców. W konsekwencji ocena wytrzymałości skorupy jaj kur mieszańców wydaje się być cechą bardzo obiecującą.
Wydajność mieszańców okazała się przydatna w przypadku oceny barwy skorupy, której przyrost dokładności między ocenami wzrastał między hodowlaną a genomową – od 26% do 40%. Wzrost dokładności oceny był niższy w przypadku masy jaja (15% – 20%) i zerowy w przypadku genomowej oceny wartości hodowlanej przeprowadzonych przy wykluciu. W odniesieniu do wytrzymałości skorupy jaj, wzrost ten występuje tylko u kur w wieku 80 tygodni, przy najniższych wskaźnikach wartości hodowlanej i genomowej (Druet i in., 2019).
Wnioski
Niezależnie od cech jakości jaj, korelacja genetyczna obserwowana między wynikami produkcyjnymi uzyskiwanymi w populacji kur czystorasowych a wynikami kur – mieszańców pokazuje, że cechy te powinny być rozważane i oceniane jako dwie różne grupy cech. W każdym przypadku ocena genomowa okazała się dokładniejsza od oceny hodowlanej. Zaobserwowano duże zróżnicowanie w zwiększaniu dokładności oceny między cechami. Ta niejednorodność może wynikać z różnic w strukturze cech. Możliwości zwiększenia dokładności ocen wartości hodowlanej i genomowej czystorasowych ptaków, wynikające z uwzględnienia wyników uzyskiwanych w populacjach mieszańców, różnią się w zależności od cechy i okresu oceny.




